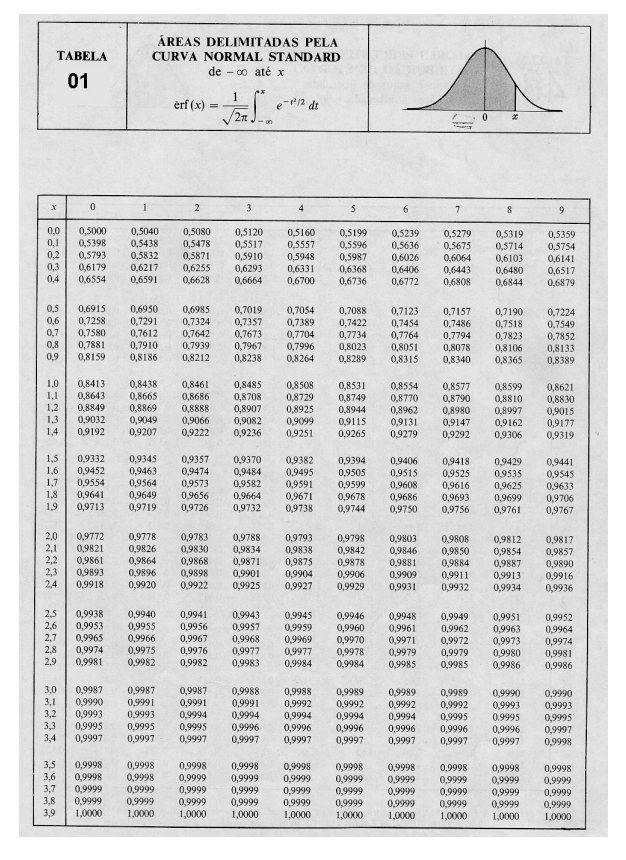
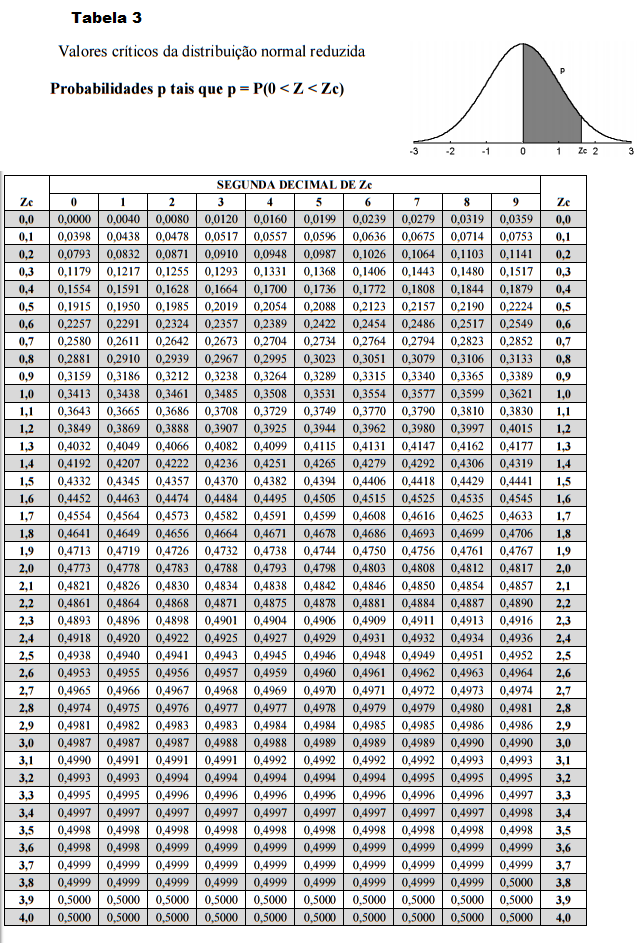
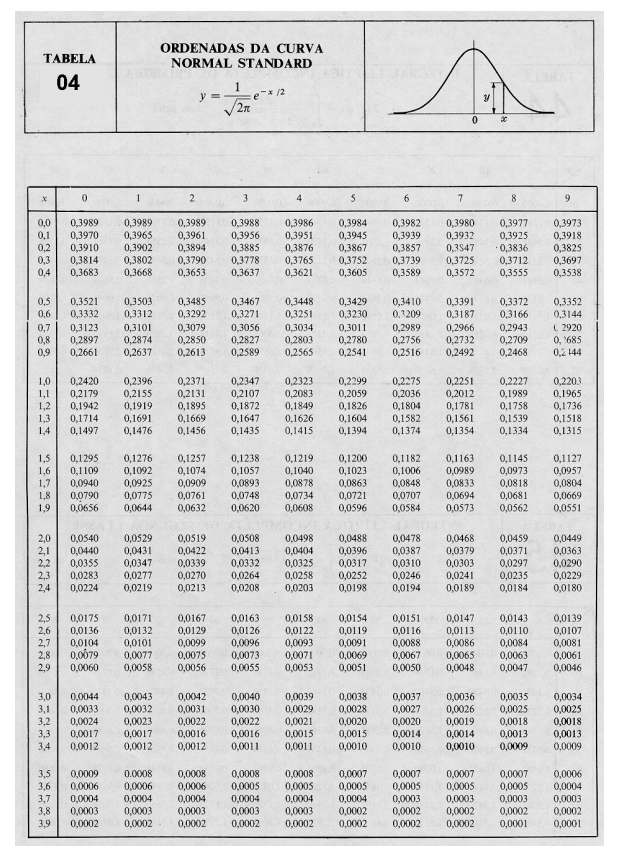
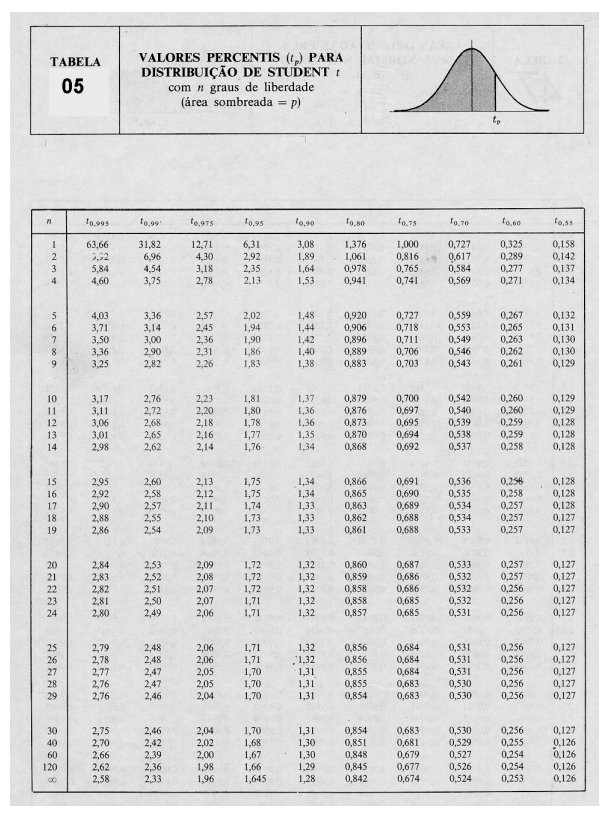
Teorias e métodos em melhoramento genético animal 4
Inicialização ao Melhoramento Genético de Produção Animal
1 Estimação de parâmetros genéticos
Os parâmetros genéticos são fundamentais para a avaliação genética dos animais e para a estruturação de programas de melhoramento genético dos rebanhos. Os parâmetros mais importantes são: herdabilidade, correlações e repetibilidade.
herdabilidade
A herdabilidade é o parâmetro que indica a consistência (confiabilidade) da utilização do desempenho fenotípico (valor fenotípico) para determinar o valor genético para determinada característica em uma população.
A herdabilidade mede também o grau de semelhança entre o desempenho fenotípico dos filhos em relação ao mesmo desempenho nos pais. Se a herdabilidade é alta, os animais com desempenho elevado tendem a produzir filhos igualmente bons e os animais com baixo desempenho tendem a produzir filhos ruins. Se a herdabilidade é baixa, o desempenho dos pais revela muito pouco sobre o desempenho da progênie.
Característica herdável e característica geneticamente determinada
Para que uma característica seja herdável é necessário que as diferenças de desempenho entre os indivíduos sejam herdáveis. Portanto, características que não apresentam diferenças fenotípicas não são herdáveis, embora sejam geneticamente determinadas. Como exemplo de característica geneticamente determinada, mas não herdável, pode-se citar o número de pernas em bovinos. Até onde se sabe, dentro da normalidade, não há diferenças entre os indivíduos quanto ao número de pernas.
Independentemente da magnitude da herdabilidade, desde que maior que zero, haverá
sempre valores genéticos altos, médios e baixos na população.
herdabilidade e semelhança entre parentes
Os procedimentos para estimação do coeficiente de herdabilidade envolvem a medição da semelhança entre parentes. Coeficiente de herdabilidade (h 2 ) é a fração da variância fenotípica que é devida às diferenças genéticas entre os indivíduos, ou seja, é a relação entre a variância genética aditiva e a variância fenotípica (h 2 = VA/VP ).
Com herdabilidade alta, os indivíduos aparentados apresentam desempenhos semelhantes, ou seja, apresentam-se semelhantes uns com os outros na característica em questão.Com herdabilidade baixa, a similaridade entre os indivíduos aparentados é
praticamente a mesma observada entre os indivíduos tomados ao acaso na população
estimação do coeficiente de herdabilidade
Como o coeficiente de herdabilidade é a razão entre a variância genética aditiva (VA) e a variância fenotípica (VP ), a estimação deste parâmetro implica na estimação dos dois componentes (VA e VP).
estimação da variância genética aditiva com base na semelhança entre parentes
Para trabalhar este conceito faz-se necessário definir dois tipos de componentes da variância fenotípica: componentes observacionais e componentes causais.
componentes observacionais da variância fenotípica(o 2 p)
São componentes de variância estimados a partir dos valores fenotípicos observados, ou seja, a partir dos dados de desempenho dos animais. Os componentes resultam da partição da variância fenotípica em partes correspondentes ao agrupamento dos indivíduos em famílias.
componentes causais da variância fenotípica (V)
São componentes da variância fenotípica, relacionados com os modos de ação gênica, ou seja, são os componentes do “modelo genético”.
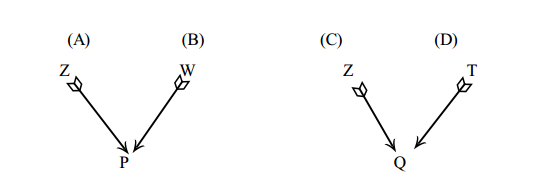
variância genética aditiva (VA)
É a parte da variância fenotípica determinada pela ação independente dos genes (ação aditiva). É a variância dos valores genéticos da população. A variância genética aditiva é
calculada com base na esperança dos componentes observacionais em termos de componentes causais da variância fenotípica.
2 Variâncias nos grupos
variância entre grupos (o 2 B)
É a variância das médias dos grupos em relação à média da população. Esta variância expressa a quantidade de variação que é comum aos membros de um mesmo grupo, o que corresponde à covariância dos grupos.
variância dentro de grupos(o 2 W)
É a variância dos valores dos indivíduos em relação à média do seu grupo. Quanto maior a similaridade (menor variação) dentro dos grupos, maior a variância entre os grupos.
correlação intraclasse
O grau de similaridade entre os indivíduos é dado, então, por

Assim, a covariância observada (covariância obtida dos dados de desempenho dos animais) é determinada por componentes que dependem do grau de parentesco. A covariância observada é calculada pelos métodos estatísticos normais (estatística básica).
Relacionando-se a covariância observada com sua esperança, em termos de componentes causais, estima-se a contribuição da variância aditiva, assim como a de outros componentes causais da variância fenotípica (dominância, por exemplo). O modelo matemático da covariância é o mesmo da variância, ou seja,
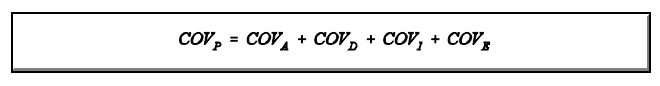
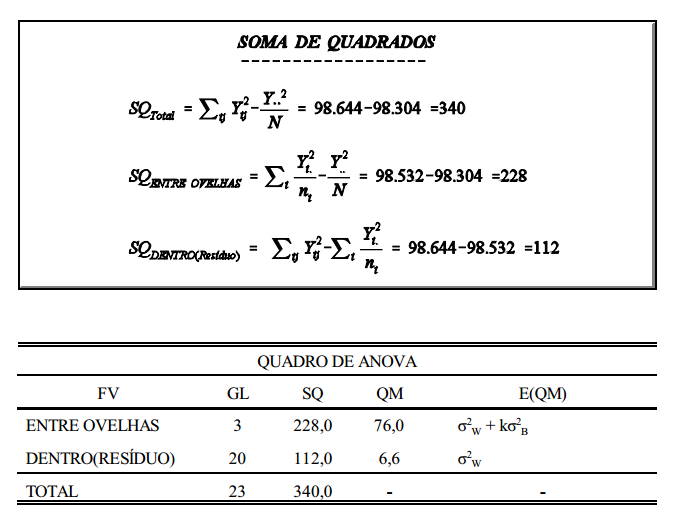
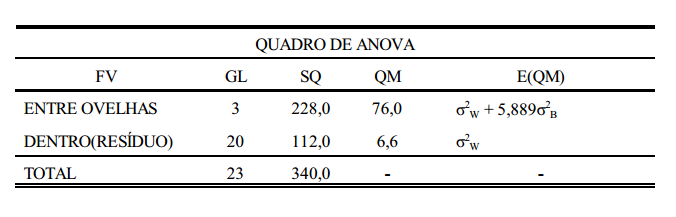
Este conceito pode ser melhor entendido com um exemplo prático, como o da tabela abaixo, que contém dados de peso de ovelhas agrupadas em famílias de irmãos completos.
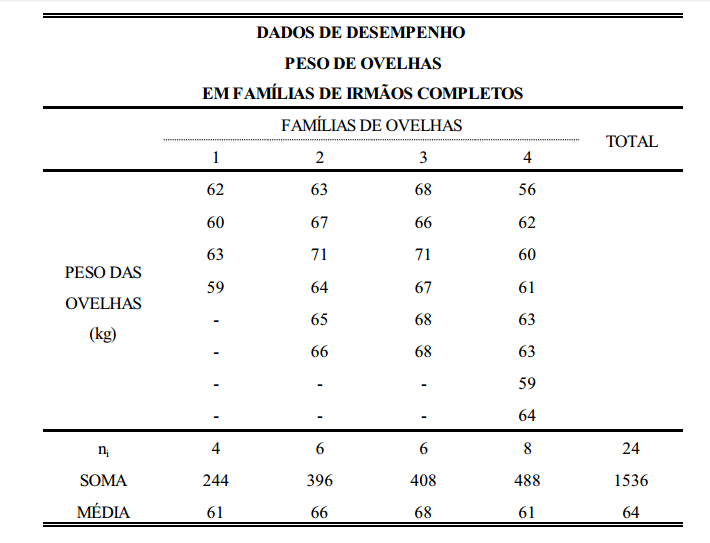
Considerando-se que a tabela acima seja o conjunto de dados em uma população de ovelhas (rebanho ou conjunto de rebanhos), a primeira coisa a ser feita, para estimação da variância aditiva, é estimar os componentes de variância entre famílias (grupos), dentro de famílias e total. O componente total, soma dos componentes estimados, é a variância fenotípica. Da mesma forma, estima-se a covariância. Para isto, utiliza-se a metodologia de análise de variância (ANOVA).
Uma vez obtidos os componentes de variância, estes serão relacionados com os componentes causais correspondentes. Como mencionado anteriormente, os componentes causais se referem às variâncias causadas pelos modos de ação gênica (aditiva, dominância e epistasia).
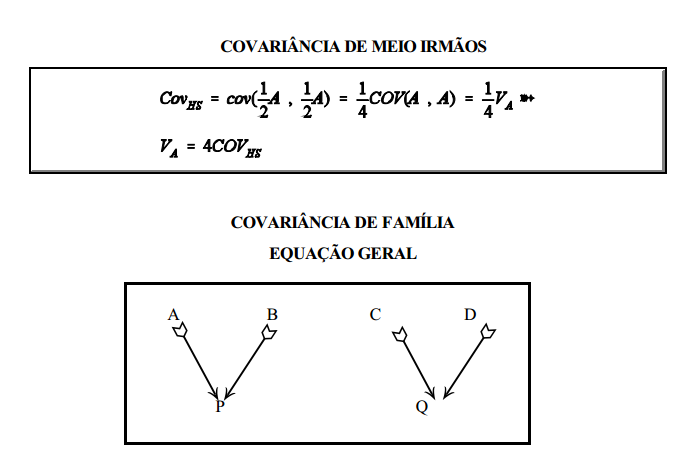
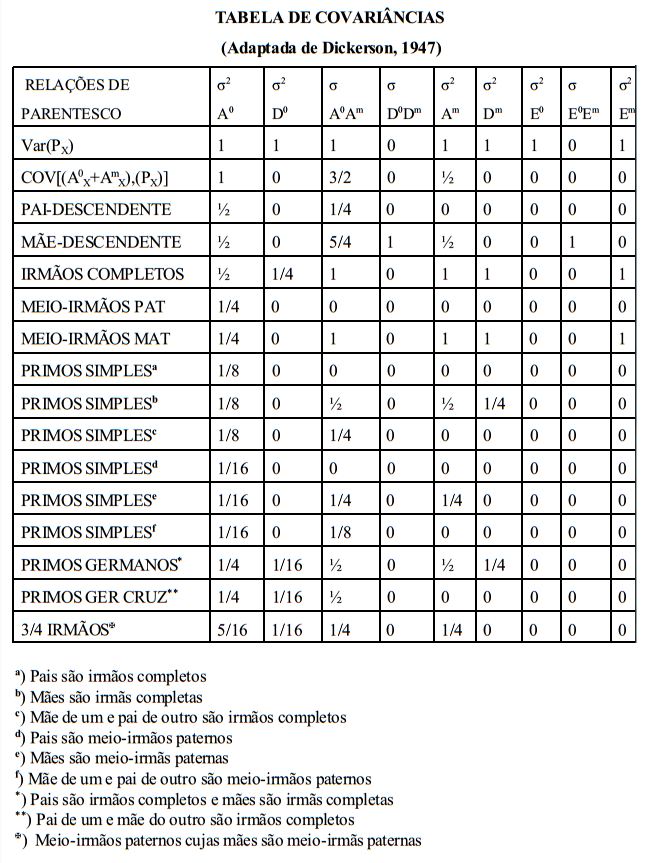
Existem algumas covariâncias entre grupos de parentes que são fundamentais (ou básicas) para a estimação da variância aditiva na população. Essas covariâncias básicas e suas esperanças em termos dos componentes causais serão trabalhas a seguir.

A covariância entre progênie e mãe deveria, pela lógica, também estimar ½ da variância aditiva. Isto não ocorre, entretanto, porque o componente observacional da covariância entre progênie e mãe envolve também a variância devida aos efeitos maternos (variância devida às diferenças de habilidade materna entre as fêmeas).

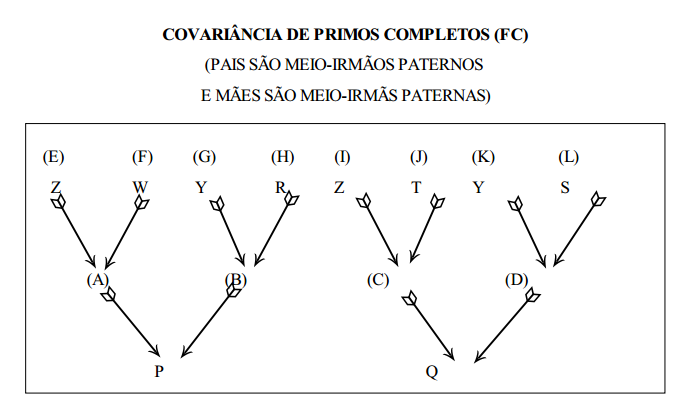
covariância de meio-irmãos paternos (PHS) usando a equação geral

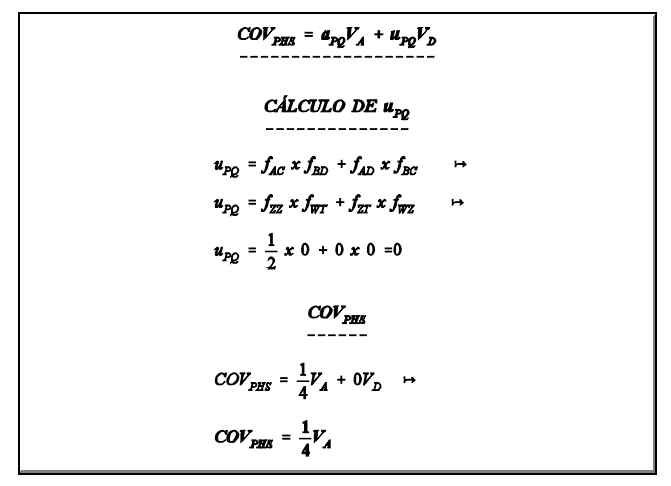
covariância de irmãos completos (FS) usando a equação geral
Sendo irmãos completos, os pais são o mesmo animal (Z) e um único animal é mãe dos dois indivíduos (W)
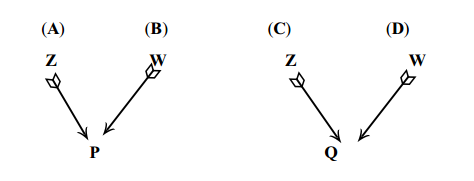

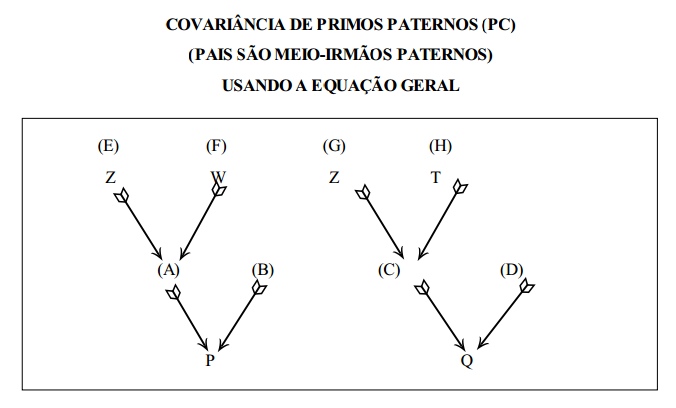

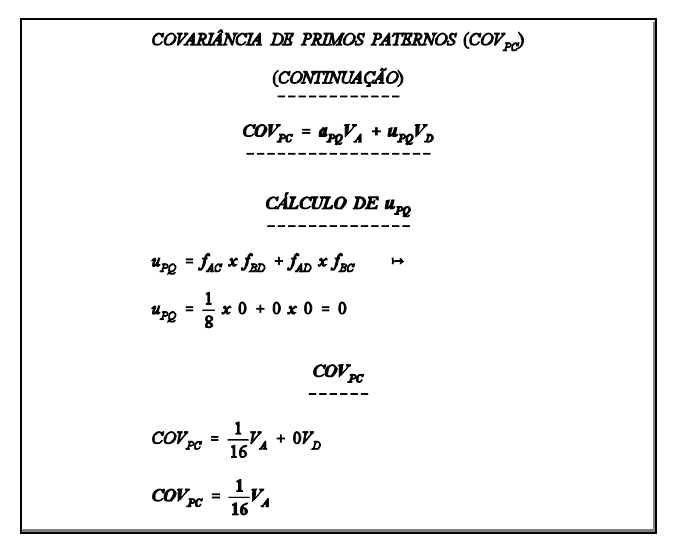


estimação da herdabilidade usando a covariância progênie X pai
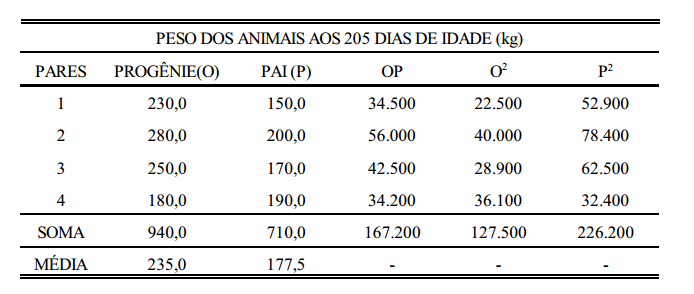

estimação da herdabilidade usando a covariância progênie X pai, a regressão progênie X pai

3 Correlações I
correlação genética ( rg )
A correlação genética entre duas características é, segundo PIRCHNER (1983), a correlação entre efeitos dos genes que as influenciam. FALCONER (1981) a define como correlação entre os valores genéticos dos indivíduos para as duas características sob consideração.
A correlação genética pode ser causada por pleiotropia ou por ligação gênica. A primeira
é própria do gene, em termos de sua capacidade de causar correlação transiente entre as características (FALCONER, 1981; PIRCHNER, 1983). A correlação transiente pode desaparecer se a ligação gênica atinge o ponto de equilíbrio.
A importância do estudo de correlações está no fato de que a seleção para uma determinada característica pode causar resposta em outra geneticamente relacionada.
correlação ambiental (re)
O coeficiente de correlação ambiental (re ) não representa exatamente a correlação devida aos fatores ambientais, mas sim uma correlação residual incluindo, além dos fatores ambientais, aqueles devidos aos efeitos de dominância, da epistasia e da interação genótipo x ambiente.
correlação fenotípica ( rp )
O coeficiente de correlação fenotípica é o coeficiente de correlação entre os valores fenotípicos dos indivíduos, para as duas características.
análise de variância e de covariância
O método mais comum para se estimarem as correlações é o da análise de meio-irmãos paternos. Pela análise de variância são estimados os componentes da variância fenotípica referentes às duas características (X e Y) cuja correlação se pretende estimar. A forma é a mesma utilizada na estimação do coeficiente de herdabilidade.
Tal como na estimação dos componentes de variância, são mostrados a seguir dois tipos de modelo básico concernentes à determinação dos componentes de covariância.
modelo 1
Um grupo de reprodutores é acasalado, cada reprodutor com d fêmeas, tendo cada fêmea apenas um filho. A forma de análise da covariância é apresentada na TABELA 1A. Se, no entanto, os reprodutores forem acasalados com números diferentes de fêmeas, a forma de análise passa a ser a da TABELA 1B.
modelo 2
Um grupo de reprodutores é acasalado, cada reprodutor com d fêmeas, tendo cada fêmea r filhos. A forma de análise é apresentada na TABELA 2A. Se o modelo for desbalanceado (diferente número de fêmeas e/ou diferente número de filhos por fêmea) a forma é a da TABELA 2B. Neste caso, os coeficientes das covariâncias na esperança dos produtos médios serão, como na análise de variância (k1, k2 e k3 ), calculados da mesma forma.

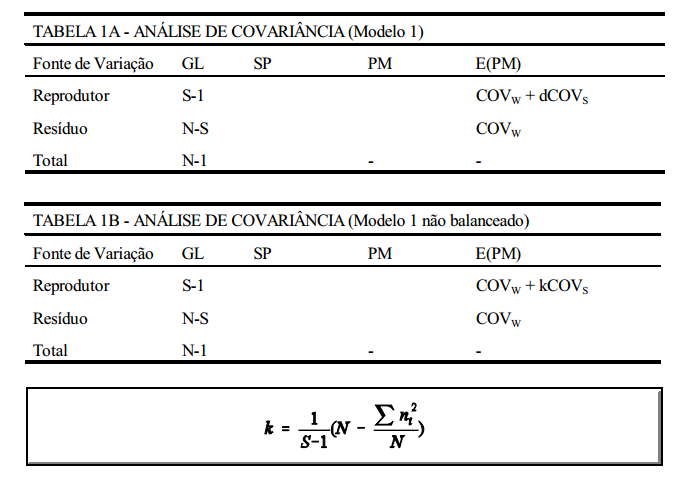
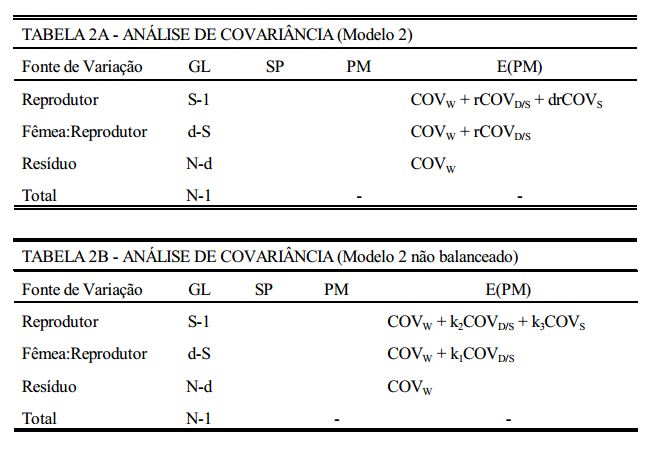
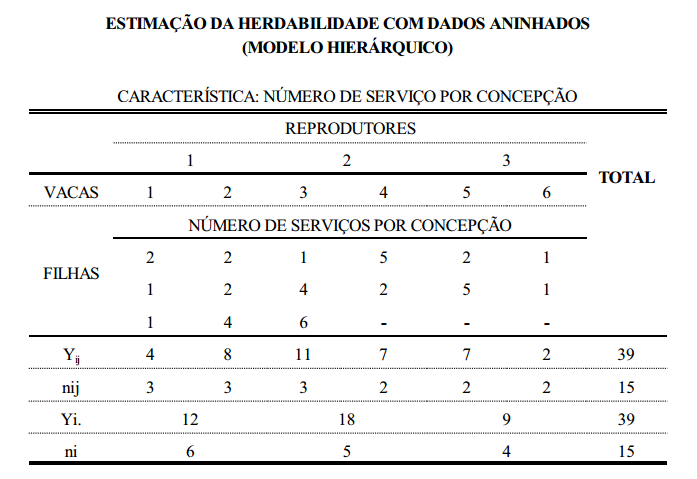
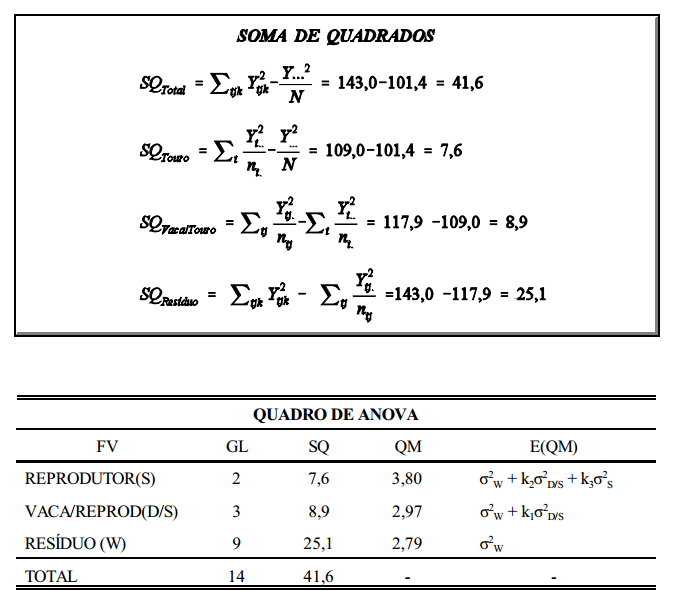
componentes da variância



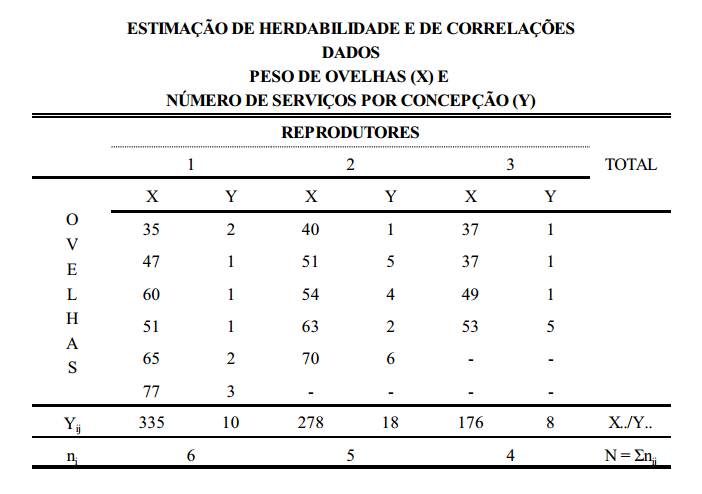


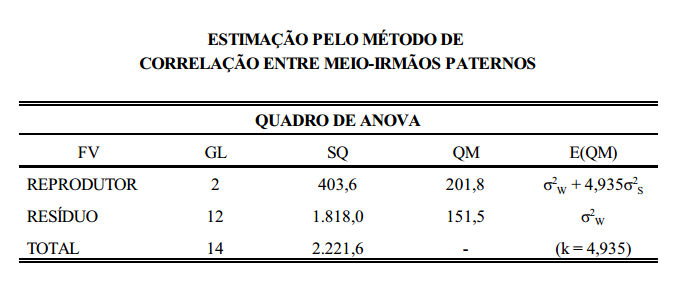


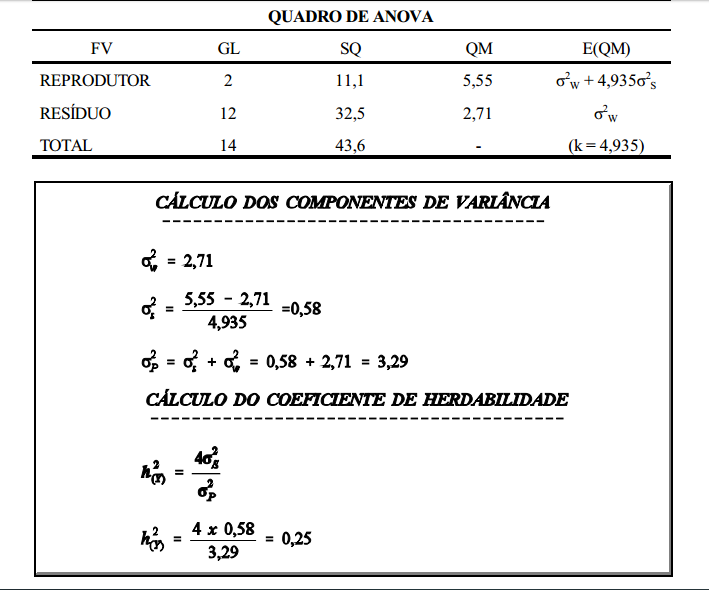
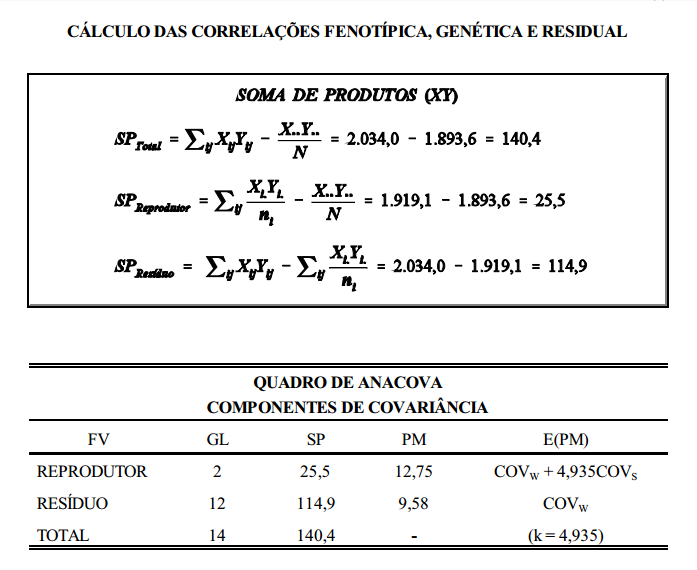

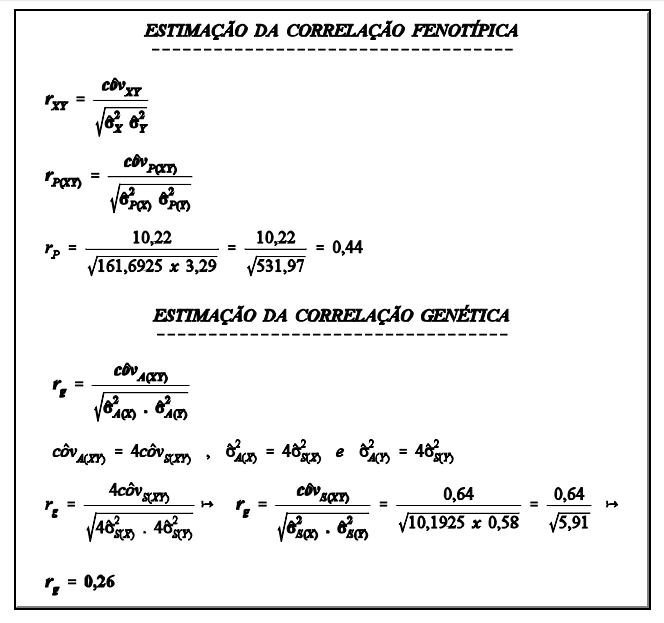
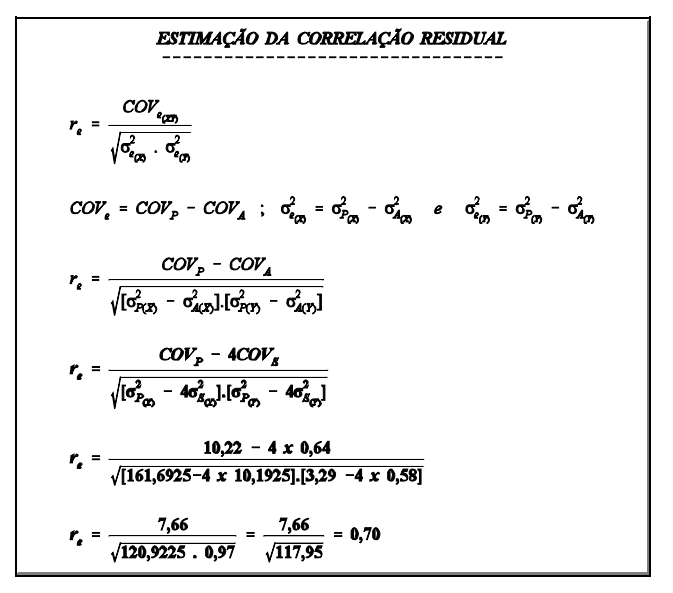
interpretação
Ovelhas mais pesadas apresentaram maior número de serviços por concepção. A correlação foi, no entanto, baixa, indicando que em muitos casos esta correspondência não existe. Em muitos casos serão encontradas ovelhas pesadas com baixo número de serviços por concepção.
Ressalta-se também que os valores utilizados neste exemplo não são reais, ou seja, a interpretação acima é apenas com relação ao exemplo e não em relação à população de ovelhas em geral. Com dados reais, o resultado poderia ter sido diferente.

4 Correlações II
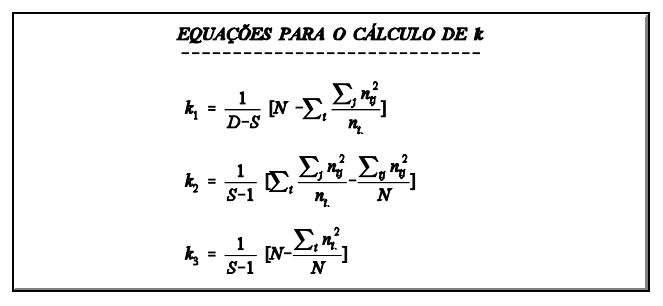

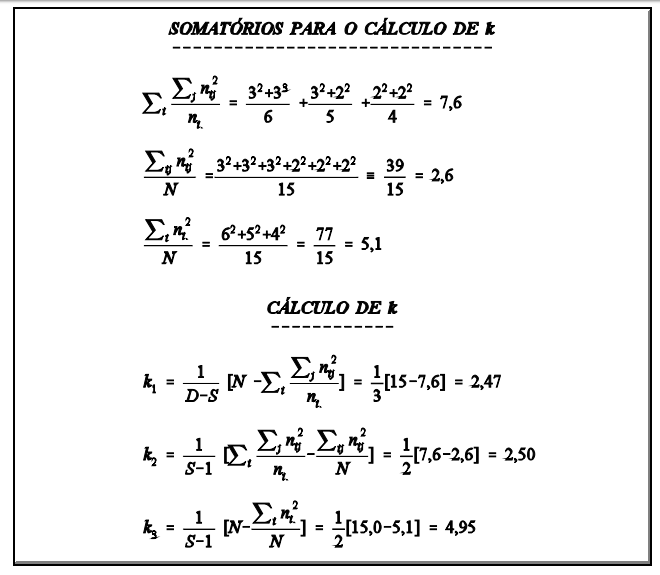

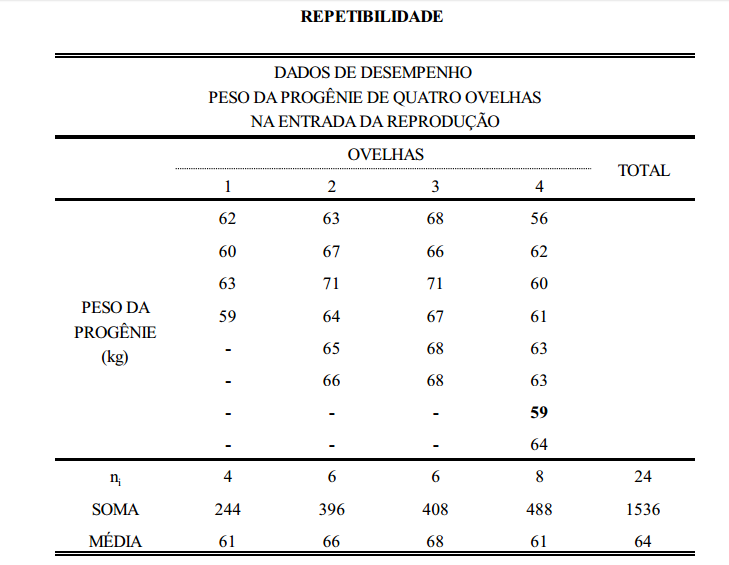
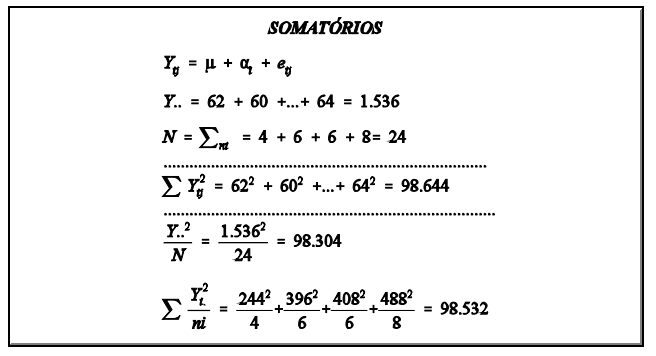


5 Efeito genético materno
conceituação
Nos mamíferos, certas características, principalmente as de crescimento, são influenciadas por dois genótipos, cada um funcionando em um indivíduo diferente e em gerações diferentes.
Ocorrem assim, duas formas de expressão gênica: uma, direta, efeito dos genes do próprio indivíduo no qual a característica é medida (efeito genético direto) e outra, indireta, efeito dos genes da mãe do indivíduo, condicionando o ambiente que esta provê a seu produto durante a gestação e na fase de aleitamento (efeito materno).
O valor fenotípico de um indivíduo é então condicionado pela expressão dos genes para crescimento herdados do pai, dos genes para crescimento herdados da mãe e dos genes para habilidade materna.
Em relação à mãe, o efeito materno resulta de seu genótipo associado a fatores ambientais, mas em relação à progênie, a influência materna pode ser considerada de natureza estritamente ambiental.
Obtenção de estimativas de coeficientes de herdabilidade
Métodos que não consideram o efeito materno podem levar a estimativas tendenciosas de coeficientes de herdabilidade. Pode ser importante selecionar vacas e touros para habilidade materna.
Correlação entre efeitos genéticos direto e materno
Vários trabalhos indicam a existência de correlação negativa entre efeitos genéticos direto e materno, o que poderia comprometer a resposta à seleção. Uma resposta positiva para potencial de crescimento poderia estar associada a uma redução na habilidade materna das vacas. As vacas com habilidade materna reduzida, produziriam bezerros mais leves.
Cruzamentos recíprocos
As mais simples evidências do efeito materno são obtidas a partir de experimentos de cruzamentos recíprocos, os quais produzem indivíduos geneticamente similares porém fenotipicamente diferentes, por serem gerados e criados por mães que diferem na habilidade materna. Exemplo: F1 de touros Nelore x vacas Angus comparados com F1 de touros Angus x vacas Nelore.
Comparação de coeficientes de herdabilidade
Os efeitos maternos podem ser detectados quando se comparam coeficientes de herdabilidade obtidos pelos métodos de correlação entre meio-irmão paternos e correlação entre meio-irmãos maternos. Também quando se comparam os coeficientes de herdabilidade obtidos pelas covariâncias entre progênie e pai e covariância entre progênie e mãe. Por exemplo, o efeito materno contribui para a correlação entre meio-irmãos maternos mas não o faz para a correlação entre meio-irmãos paternos. A diferença entre as duas correlações seria teoricamente, imputada ao efeito materno e à covariância entre potencial de crescimento (efeito genético direto) e efeito materno.
Modelo de Dickerson
Dickerson (1947) mostrou com diagramas de passagem (figura 1) a influência dos efeitos genéticos direto e materno sobre o valor fenotípico e determinou a correlação genética entre os dois efeitos.
Modelo de Wilham
Wilham (1963) desenvolveu um modelo baseado na composição teórica esperada para a covariância entre indivíduos com diferentes graus de parentesco. Este modelo, para uma característica sujeita à influência materna, considera o valor fenotípico de um indivíduo X qualquer, de mãe W, como sendo resultante da ação do fenótipo de X (P O
X ) e da ação do fenótipo materno (P m
W), defindo "o" como componente do próprio indivíduo cuja característica
é medida (efeito direto) e "m"como componente materno (efeito materno), ou seja,





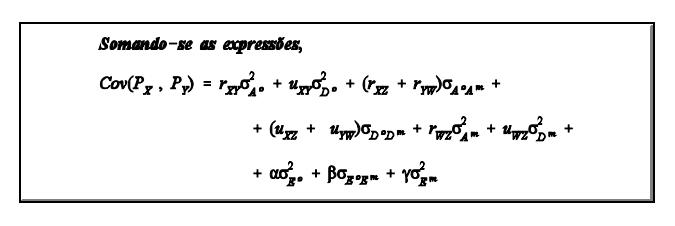

6 Estimação dos componentes causais usando dados de campo
Com este modelo (WILHAM, 1963) a estimação dos componentes causais da variância fenotípica segue os seguintes passos:
1. Estimar, dos dados, os componentes de variância e de covariância para todas as relações de parentesco (famílias ) disponíveis ( meio-irmãos paternos, meio-irmãos maternos, primos, covariância entre progênie e mãe etc. Usar GLM dos SAS, por exemplo).
2. Igualar cada componente à sua composição teórica (vide Tabela 1). Forma-se um sistema de n equações e p incógnitas, em que n é o número de relações de parentesco disponíveis e p é igual ao número de componentes causais de variância e de covariância.
3. Resolver o sistema de equações formado. Se n for <> de p pode-se aplicar o método dos quadrados mínimos (como proposto por EISEN, 1967).
4. O componente de variância fenotípica (o 2 P ) será igual à soma das estimativas dos componentes causais.
5. A relação entre o valor estimado para o componente genético aditivo direto e a variância fenotípica fornecerá uma estimativa da herdabilidade para o efeito direto. Para se estimar a herdabilidade do efeito materno, a relação será entre o componente genético aditivo materno e a variância fenotípica.
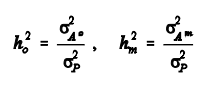


exemplo com dados de campo
Um exemplo completo de estimação do efeito materno, utilizando a metodologia descrita acima, pode ser obtido em Eler 1987.
7 Probabilidade de detecção de portadores de genes recessivos
É importante saber se um reprodutor (ou uma matriz) com fenótipo dominante é
homozigoto para o gene dominante ou se é heterozigoto. É importante sobretudo saber se o homozigoto recessivo apresenta uma característica indesejável.
Não há jamais completa certeza de que um animal não seja portador de recessivos; há
todavia métodos que permitem assegurar com certa probabilidade.
Entre os métodos de detecção podem ser citados:
informações do “Pedigree”
a) O animal é conhecidamente portador se um dos pais tem genótipo homozigoto recessivo.
b) Progênie de portadores são provavelmente portadores.
No que concerne aos filhos de machos que são sabidamente heterozigotos para genes recessivos indesejáveis, duas alternativas podem ser utilizadas:
• Retirá-los sistematicamente da reprodução.
• Testá-los para identificar os heterozigotos.
teste de progênie para identificação dos heterozigotos
Há várias alternativas para o teste de progênie do macho, dependendo do tipo de fêmea disponível para acasalamento. Todas as alternativas seguem o seguinte procedimento:
1 - Assumir que o animal suspeito seja de fato heterozigoto.
2 - Calcular a probabilidade (p) de se observar o fenótipo dominante em uma única progênie.
3 - Calcular a probabilidade de n progênies observadas mostrarem o fenótipo dominante (p n).
4 - Calcular a probabilidade de que pelo menos 1 fenótipo recessivo possa ser observado. Esta probabilidade é dada por (1-p n).
A fração (1-p n ) é, portanto, a probabilidade de detecção do gene recessivo.
8 Alternativas na utilização do teste de progênie
a) Uso de fêmeas homozigotas recessivas
A probabilidade de uma progênie mostrar o fenótipo dominante, do acasalamento de Aa com aa é 1/2. p=1/2. A probabilidade de n dominantes é p n = (1/2) n. A probabilidade de pelo menos um apresentar o fenótipo recessivo é 1-(1/2) n. Desta forma, tem-se:


b) Uso de fêmeas portadoras


c) Uso de fêmeas filhas de um touro portador f (A) = 3/4 e f(a) = 1/4


d) Uso de fêmeas filhas do suspeito

É idêntico ao anterior, portanto a probabilidade de detecção é igual a 1 - (7/8) n e o número de progênies necessário para 95 % de confiança é 23 e para 99% é 35.

e) Acasalamentos aleatórios




Conclui-se, assim, que, para frequências muito baixas do recessivo, a utilização de acasalamentos aleatórios não permite, na prática, níveis elevados de confiabilidade para o reprodutor. A obtenção de 598 progênies do reprodutor, por exemplo, seria muito difícil e os custos poderiam ser elevados.
9 Bibliografia recomendada
BOURDON, R.M. Understandig Animal Breeding. Prentice Hall, 1997.
CHAPMAN, A..B. General and Quantitative Genetics. New York, Elsevier, 1985. 408p.
CROW, J.F. & KIMURA, M. An Introduction to Population Genetics Theory. New York, Harper & Row, 1979, 591p.
DALY, J.J. Melhoramento Genético para Produção de Carne Bovina. Tradução: Gensys. Edição: Agropecuária CFM Ltda., 1977.
DICKERSON, G.E. Composition of hog carcass as influenced by heritable diferences in rate and economy of gain. Iowa Agr. Exp. Sta. Bul. 354, 1947.
ELANDT-JOHNSON, R.C. Probability Models and Statistical Methods in Genetics. New York, Wiley, 1971.
ELER, J.P. Teorias e Métodos em Melhoramento Genético Animal. Apostila, 1999, 230p.
FALCONER, D.S. Introduction to Quantitative Genetic. London, Longman Group Corp., 1981, 340p.
FOULEY, J.P. & LEFORT,G. Méthodes d’estimation des effectes directs et maternels em sélection animale. Ann. Génet. Sél. Anim., 10(3): 475-496, 1978.
GIANONI, M.A. & GIANONI, M.L. Genética e Melhoramento de Rebanhos nos Trópicos. São Paulo, Nobel, 1983, 460p.
HENDERSON, C.R. Seletion index and expected genetic advance. In Statistical Genetics and Plant Breeding. NAS-NRC 982: 141-163, 1963.
JOHANSON, I & RENDEL, J. Genética Y Meyora Animal. Zaragoza, Acribia, 1972.
KEMPTHORN, O. An Introduction to Genetic Statistics. New York, Wiley, 1957.
LASLEY, J.F. Genetics of Livestock Improvement. New Jersey, Prentice Hall, 1978, 492p.
LERNER, I.M. & DONALD, H.P. Recentes Progressos no Melhoramento Genético dos Animais. São Paulo, EDUSP, 1969.
LI, C.C.. First Course in Population Genetics. Pacific Grove (California), The Boxwood Press, 1976. 631p.
LUSH, J.L. Melhoramento Genético dos Animais Domésticos. Rio de Janeiro, USAID.
PEREIRA, J.C.C. Melhoramento Genético Aplicado à Produção Animal. FEP-MVZ Editora, Belo Horizonte, 1999.
PIRCHNER, F. Population Genetics in Animal Breeding. London, Plenum Press, 1983.
SEARLE, R.; CASELA,G. AND McCULLOCH, E. Variance Components. John Wiley & Sons, INC.
SILVA, R.G. Métodos de Genética Quantitativa Aplicados ao Melhoramento Genético Animal. Ribeirão Preto, Sociedade Brasileira de Genética, 1982, 162p.
STRICKBERGER, M.W. Genetics. New York, Macmilian Publishing Company, 1985, 842p.
VAN VLECK, L.D., POLLAK, E.J. & OLTENACU, E.A.B. Genetics for the Animal Sciences, New York, W.H. Freeman and Company, 1987, 391p.
VANVLECK,L.D. Selection Index and Introduction to Mixed Models Methods. CRC Press, Boca Raton, FL, 1993.
WARWICK, E.J. & LEGATES,J.E. Breeding and Improvement of Farm Animals. New York, McGrow Hill, 624p.
WILLHAM, R.L. The covariance between relatives for characters composed of components contributed by related individuals. Biometrics, 19: 18-26, 1963.
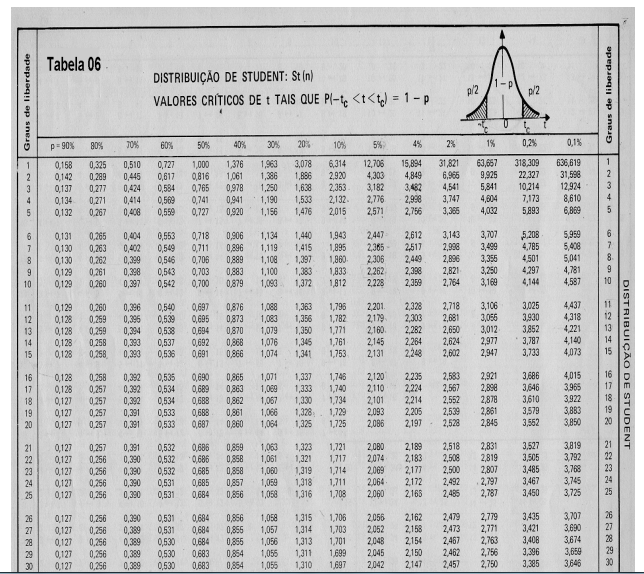
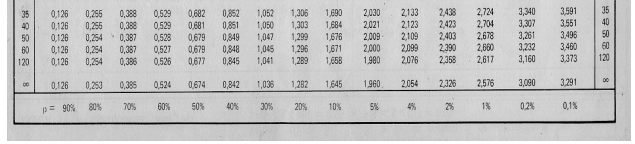
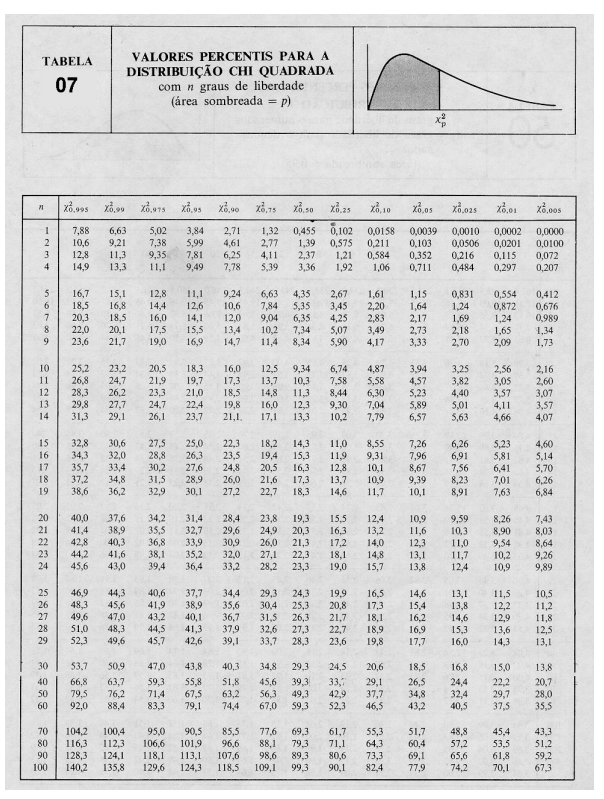
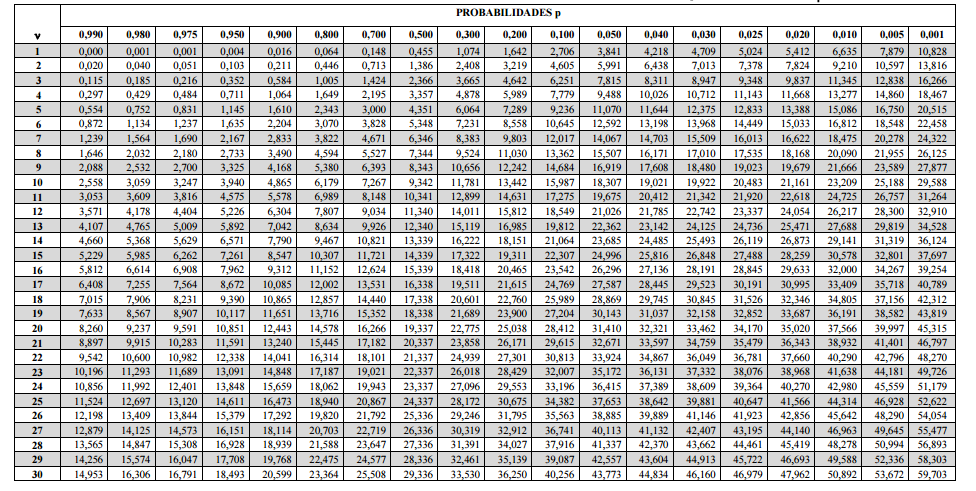
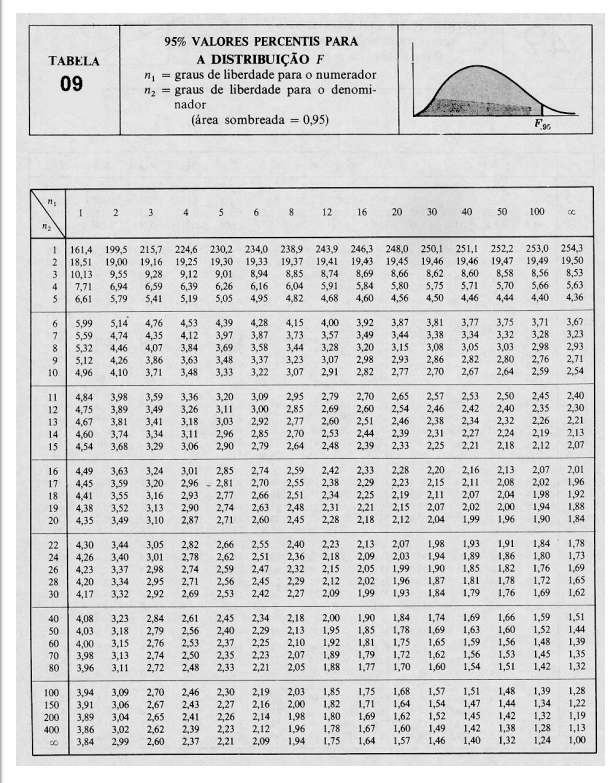
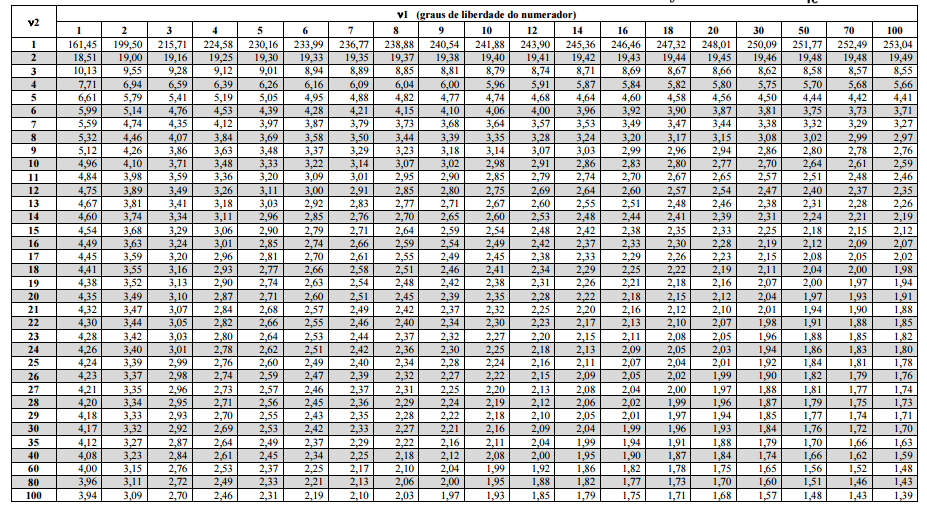
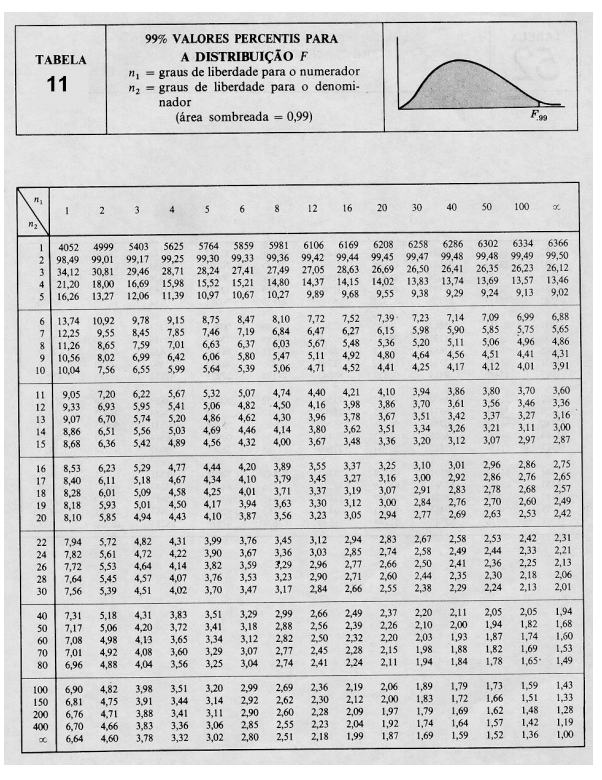
10 Tabelas