Teorias e métodos em melhoramento genético animal 2
Inicialização ao Melhoramento Genético de Produção Animal
1 Introdução à genética de populações I
Conceito de população
a) do ponto de vista estatístico ---> é o total de indivíduos de uma espécie ou raça ou de outro grupamento qualquer que habitam determinada área.
b) do ponto de vista genético ---> é um grupo de indivíduos da mesma espécie, coexistindo em uma área comum e reproduzindo-se. Esta categoria de população recebe o nome de População Mendeliana.
Genética de populações
É o ramo da ciência que estuda as frequências gênicas e genotípicas nas populações e as forças capazes de alterá-las ao longo das gerações.
Entende-se por frequência gênica, a proporção dos diferentes alelos de um gene na população, enquanto que frequência genotípica é a proporção dos genótipos.
Cálculo de frequências gênicas
O cálculo de frequências gênicas depara com duas situações distintas: ausência de dominância e dominância completa.
Neste caso, considerando-se um locus com dois alelos, têm-se três fenótipos distintos e a frequência de determinado alelo é dada por:

Exemplo: na herança da pelagem do gado Shorthorn não existe dominância, ou seja, o gene A1 em homozigose produz a cor vermelha e o gene A2 em homozigose produz a cor branca. Quando ocorre heterozigose, os indivíduos são ruões. Em um rebanho de 100 animais, onde 50 são vermelhos, 40 ruões e 10 brancos, qual é a frequência dos genes A1 e A2?

A probabilidade de qualquer gameta transportar um determinado gene é igual à frequência desse gene na população. A probabilidade de um espermatozoide portador do gene A1 fertilizar um óvulo A1 é, nesse caso, 0,7 x 0,7 = 0,49.
b) Dominância completa
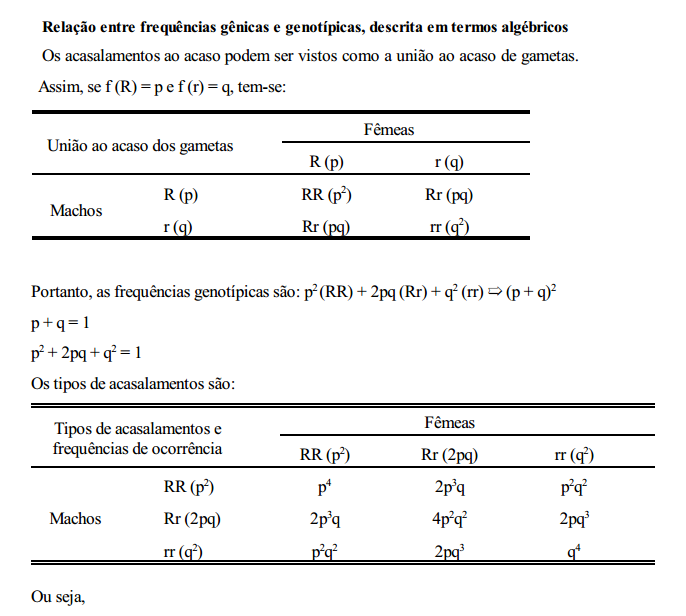
Neste caso, têm-se três genótipos e apenas dois fenótipos distintos e, para o cálculo das frequências gênicas, torna-se necessário pressupor que a população se encontra no equilíbrio de Hardy-Wenberg.
Equilíbrio de Hardy-Wenberg
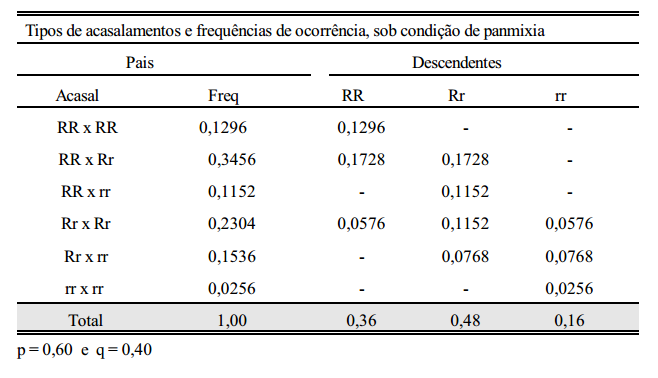
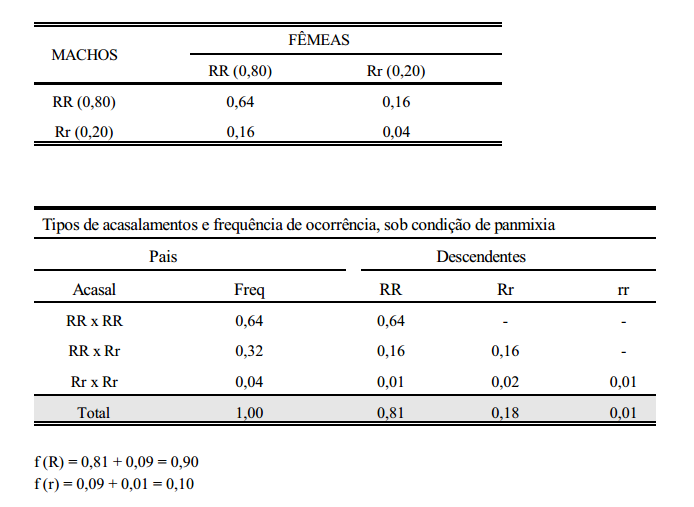
Considere-se como característica de estudo, a pelagem de uma certa população de suínos, em que os alelos R e r controlam a sua expressão. Os indivíduos homozigotos dominantes (RR) e os heterozigotos (Rr) são vermelhos e os homozigotos recessivos (rr) são pretos. Partindo-se de uma população inicial composta de um número arbitrário de cada genótipo, pode-se prever as frequências gênicas e genotípicas nas gerações que se seguem. considere-se, por exemplo, a geração 0 (zero) composta de 40% de RR, 40% de Rr e 20% de rr, ou seja,

As frequências gênicas seriam dadas por:
f (R) = p = 0,40 + 0,20 = 0,60
f (r) = q = 0,20 + 0,20 = 0,40
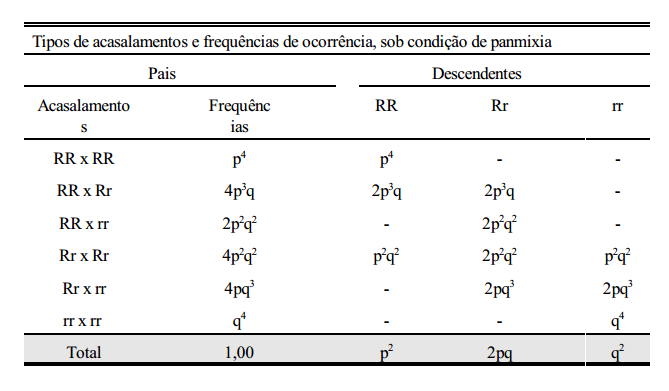
Considerando-se que a cor da pelagem não interfira nos acasalamentos, ou seja, que estes sejam feitos ao acaso, os seguintes tipos de acasalamentos com suas respectivas frequências de ocorrências são apresentados abaixo.
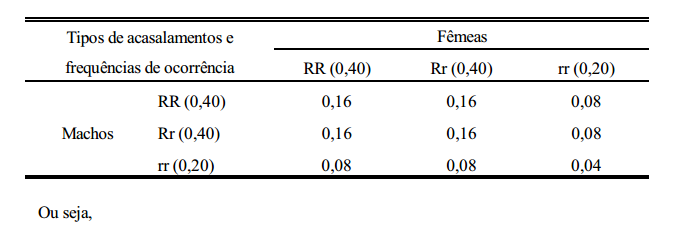

Do que se conclui que:
1. Sob condição de panmixia (acasalamentos ao acaso) em uma população de grande efetivo, a frequência gênica em determinada geração depende da frequência gênica da geração anterior e não da frequência genotípica.
2. A frequência genotípica em determinada geração depende da frequência gênica da geração anterior.
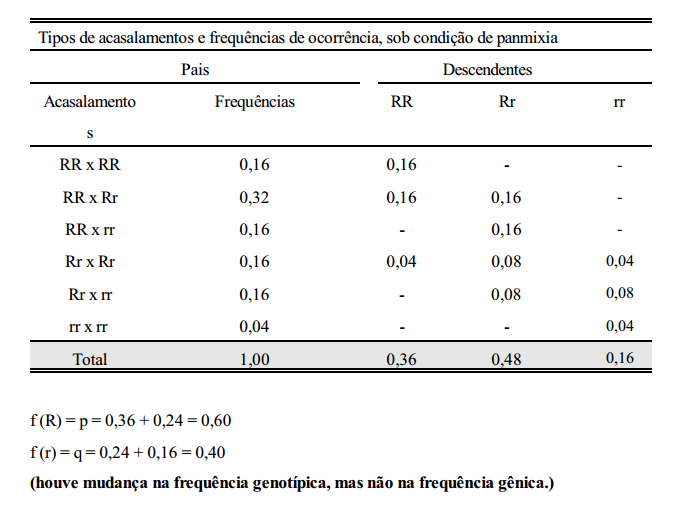
Considere-se agora, a seguinte população: 36% RR, 48% Rr e 16% rr.
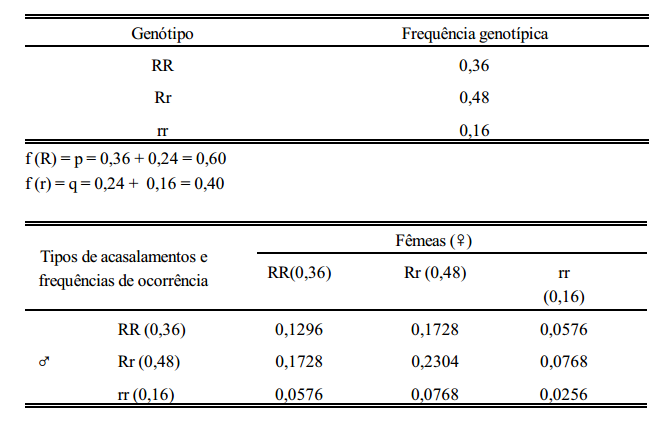
conclusão
Em uma população panmítica grande, as frequências gênicas e genotípicas permanecerão constantes de geração a geração, na ausência de seleção, migração e mutação.
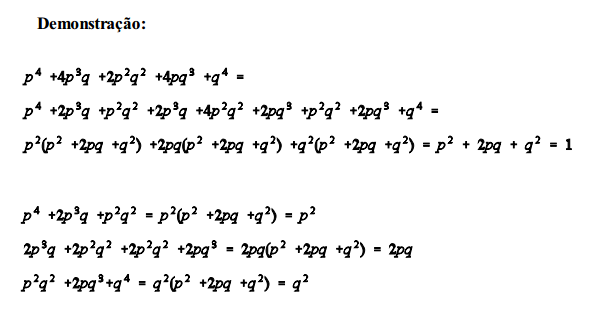
Corolário de Wentworth-Remick
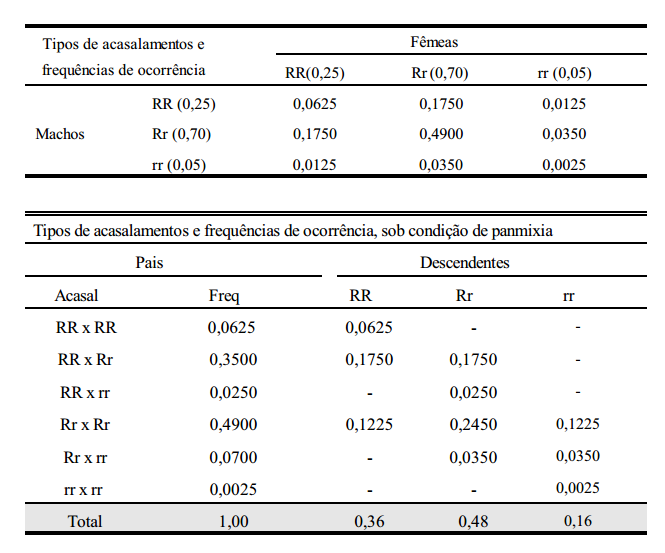
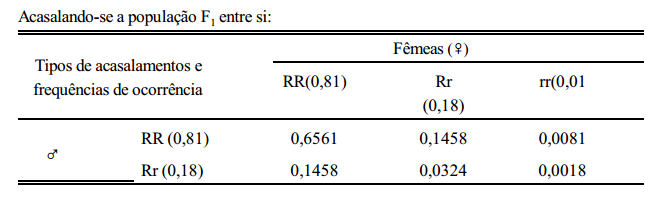
Uma população que não se encontra no equilíbrio, alcança tal estado em apenas uma geração de acasalamentos ao acaso. Considere-se a seguinte população:
RR = 80 indivíduos e Rr = 20 indivíduos.
Ou seja, f (RR) = 0,80; f (Rr) = 0,20; f (rr) = 0,00 ===> f (R) = 0,9 e f (r) = 0,1 A falta do genótipo rr caracteriza o não equilíbrio da população. Realizando-se uma geração de acasalamentos ao acaso, tem-se:
2 Introdução à genética de populações II
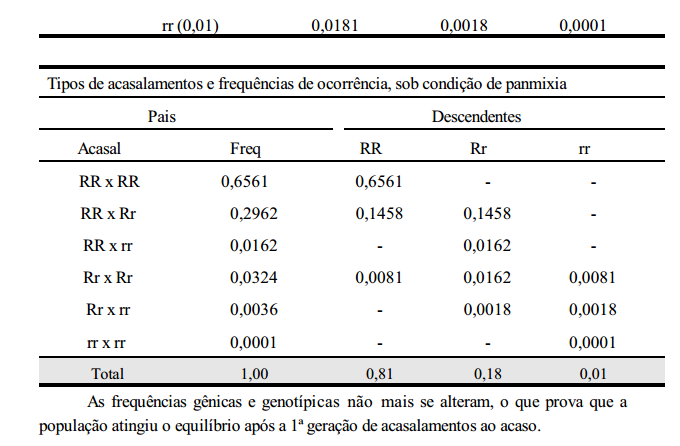
As frequências gênicas e genotípicas não mais se alteram, o que prova que a população atingiu o equilíbrio após a 1ª geração de acasalamentos ao acaso.
Calculo de frequências gênicas quando ocorre dominância completa
Neste caso, assume-se que a população está em equilíbrio. Exemplo: Em um rebanho bovino de 100 animais, verificou-se que 9 eram chifrudos e 91 mochos. Quais são as frequências dos alelos "M" e "m" na população?
Solução: MM + Mm = 91
mm = 9 ===> f (mm) = q 2 = 9/100 = 0,09
Pela suposição de equilíbrio, as frequências genotípicas esperadas são:
p 2
(MM); 2pq (Mm) e q 2
(mm). Assim,

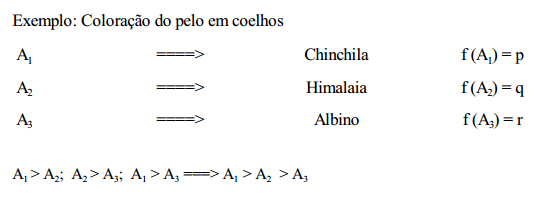
3 Alelos múltiplos
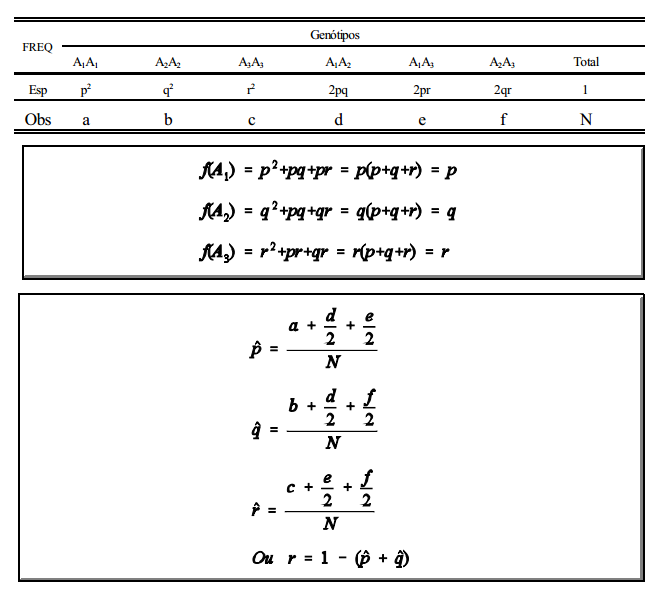
Trata-se de mais de dois alelos em um mesmo locus. Considerando-se, por exemplo, três alelos, A1 , A2 e A3 , com as frequências gênicas f (A1 ) = p; f (A2 ) = q e f A1 ) = r (com p + q + r =1), tem-se, de acordo com o teorema de Hardy-Wenberg, as seguintes frequências genotípicas no equilíbrio: p 2
(A1A1 p + q 2
(A2A2 p + q 2
(A3A3 ) + 2pq (A1A1 ) + 2pr (A1A3 ) + 2qr (A2A3 ), o que corresponde ao desenvolvimento do trinômio p + q + r 2
.
Com k alelos têm-se, então:

Estimação das frequências alélicas usando dados de amostra quando todos os fenótipos são distinguíveis
Se todos os fenótipos são distinguíveis, somam-se a frequência do homozigoto mais a metade das frequências dos heterozigotos que contenham o alelo em questão.
Estimativas de frequências alélicas usando dados de amostras quando ocorre dominância
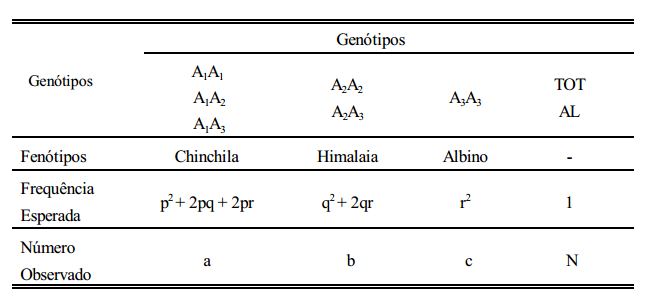
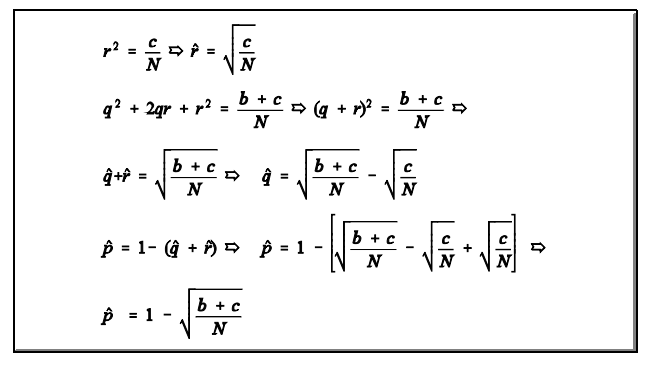
Estimativa das frequências alélicas quando ocorre dominância e codominância

Genótipos(genót), Fenótipos (Fenót),Frequências genotípicas esperadas (FrEsp) e número de observações (Núm) na estimação de frequências gênicas para grupos sanguíneos*
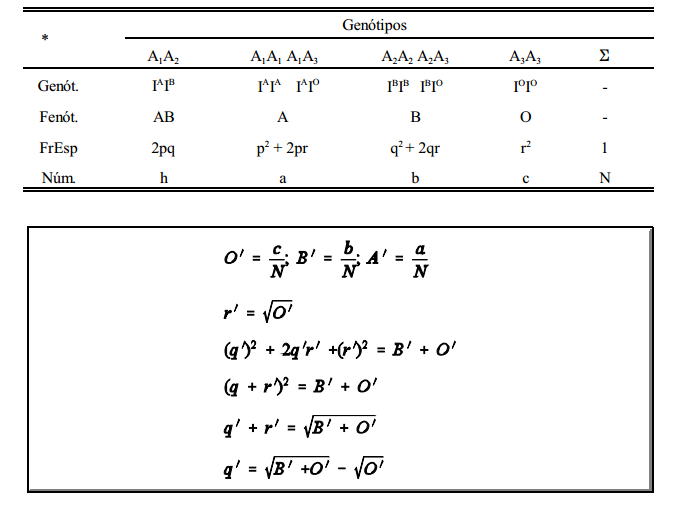
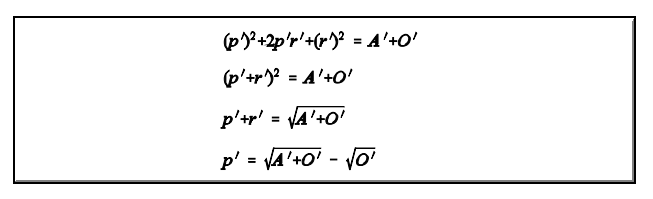

Entretanto, por estas estimativas, p' + q' + r'
1. Isto faz com que seja necessário um ajuste. As estimativas ajustadas segundo o modelo de BERNSTEIN (1925) são obtidas da forma abaixo:
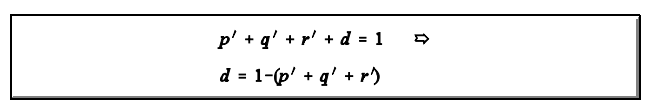
E as frequências ajustadas são dadas por:
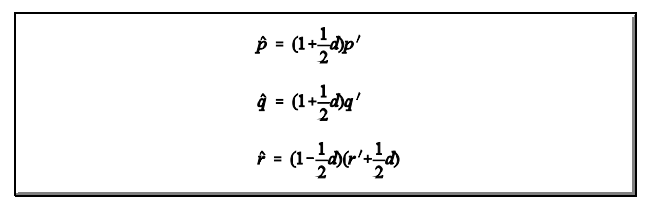
Propriedades de uma população em equilíbrio de Hardy-Wenberg com alelos múltiplos
1. O número de heterozigotos pode exceder a 50%
2. A proporção máxima de heterozigotos é 2/3 e ocorre quando p = q = r = 1/3

Para que uma função tenha máximo ou mínimo, sua derivada de primeira ordem deve ser nula. Então:

4 Genes ligados ao sexo
Introdução
Genes ligados ao sexo são genes localizados nos cromossomos sexuais. Quando o gene está ligado ao cromossoma X, a proporção genotípica no sexo que possui tal cromossoma em dose simples (sexo heterogamético - XY) será igual à
frequência do gene. Já no sexo homogamético (XX), a proporção genotípica será a mesma de um gene autossômico. Uma característica cuja expressão depende do gene recessivo ligado ao cromossomo X, terá frequência genotípica no macho igual a q e na fêmea igual a q 2.
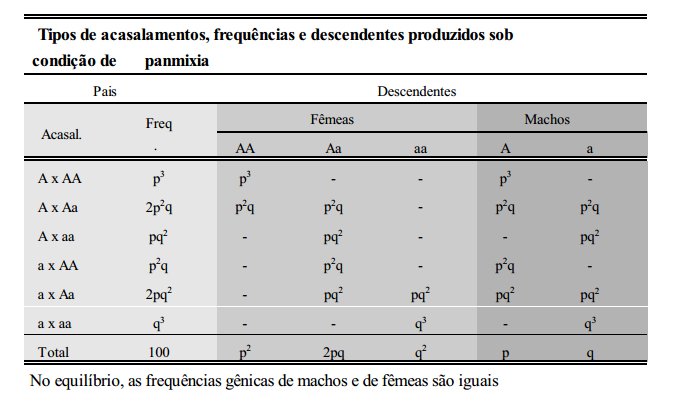
População em equilíbrio
Pode-se demonstrar que a população abaixo, sob condição de panmixia, encontra-se em equilíbrio de Hardy-Wenberg, para uma característica cuja expressão esteja relacionada aos genes A e a:

Estabelecimento do equilíbrio
Devido ao complemento cromossomal assimétrico de machos e de fêmeas, o estado de equilíbrio não se estabelece em uma única geração de acasalamentos ao acaso. Partindo-se de uma população inicial arbitrária, a condição de equilíbrio aproxima-se rapidamente, de forma oscilatória. Imagine-se a seguinte população inicial:
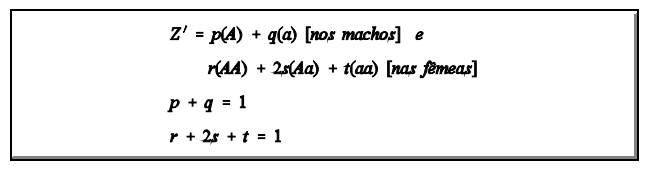
As frequências gênicas são:
f (A) = p e f (a) = q [nos machos]
f (A) = (r + s) e f (a) = (s + t) [nas fêmeas]
No cálculo das proporções dos vários genótipos da próxima geração, faz-se uso do princípio de que acasalamento ao acaso é equivalente à união aleatória dos gametas. A fêmea produz dois tipos de óvulos: (r + s) A e (s + t) a. Quando esses óvulos se unem aos espermatozoides contendo o cromossomo Y, formam os descendentes machos. Quando se unem aos espermatozoides contendo o cromossomo X, formam as fêmeas.

Frequências gênicas
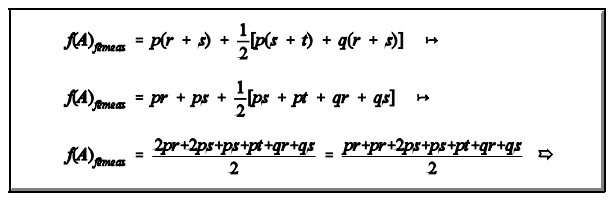

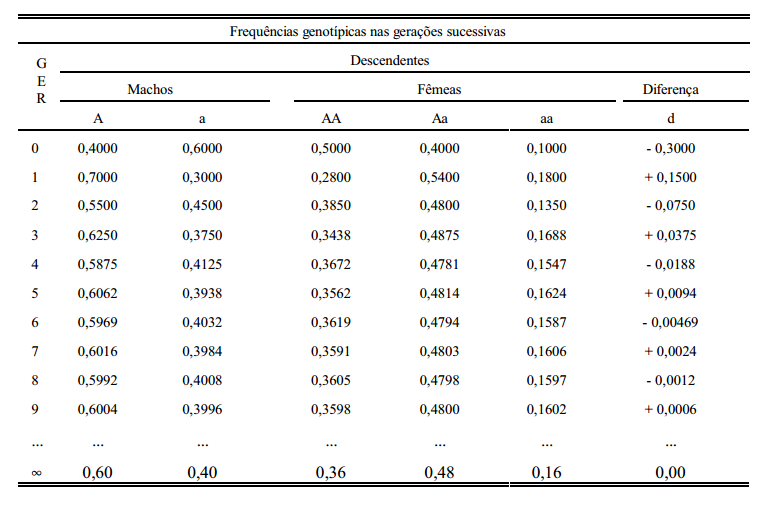

A diferença entre frequências gênicas de fêmeas e de machos se reduz à metade em cada geração (d = - 1/2 d' ).
Cálculo de frequências gênicas com genes ligados ao sexo


5 Fatores que afetam a frequência gênica
introdução
Como foi visto nos capítulos anteriores, uma população grande que mantém um sistema de acasalamentos ao acaso permanece estável com relação às frequências gênicas e genotípicas na ausência de forças externas capazes de mudar as suas propriedades genéticas. Esse estado de equilíbrio pode então ser modificado pela atuação de forças externas.
Os processos de alteração da frequência gênica podem ocorrer de duas formas:
Sistemática -Tende a alterar a frequência gênica de uma forma previsível em quantidade e em direção. São três os processos sistemáticos: migração, mutação e seleção.
Dispersiva - Ocorre em pequenas populações, como resultado de efeitos amostrais. Esta forma é conhecida como oscilação genética. Pode-se prever a quantidade da mudança, mas não a sua direção.
6 Fatores capazes de alterar as frequências gênicas I
migração
conceituação
É o movimento de indivíduos de uma população para outra, seguido de reprodução entre as subpopulações, resultando na “mistura” dos patrimônios genéticos dessas subpopulações.
No contexto da zootecnia, os exemplos mais pertinentes são a introdução do gado zebu no Brasil, com alteração da frequência de muitos genes, e a introdução das raças Simental, Charolesa e outras nos Estados Unidos, com absorção das raças locais, resultando em enormes alterações no tamanho, no ganho de peso, na produção de leite e em outras características de interesse econômico.
variação na frequência gênica
Suponha-se que uma grande população consista da proporção “m” de imigrantes em cada geração, sendo o restante, “1- m”, de nativos. Se a frequência de um gene for qm entre os imigrantes e q0 entre os nativos, a frequência do gene na população mista será q1 dado por:

conceituação
Mutação gênica é uma mudança na sequência de bases nitrogenadas do DNA de um cormossomo, com consequente mudança na síntese de RNA, que leva as informações para a síntese proteica que ocorre nos ribossomos. Desta forma, a nova proteína funcionais e, consequentemente, fenotípicas de grande importância.
As mutações podem ser de dois tipos:
Mutações recorrentes 6 mutações que ocorrem com determinada frequência.
Mutações não recorrentes 6 aquelas que ocorrem apenas uma vez e não mais se repetem.
As mutações podem ainda ocorrer nos cromossomos autossômicos ou nos sexuais,disto dependendo a sua transmissão ou não aos descendentes. Por exemplo, alguns bovinos na raça Hereford nascem com pintas pretas no corpo (animais vermelhos) que não são transmitidas. Provavelmente tais mutações ocorrem nas células embrionárias que deram origem às células somáticas, não atingindo as germinativas.
Na maioria dos casos, as mutações são indesejáveis, causando erros metabólicos que podem inviabilizar a sobrevivência do seu portador.
A frequência das mutações naturais é extremamente baixa (da ordem de 10-5, ou seja, ocorre um mutante em 100.000 indivíduos normais), entretanto as mutações têm sido importante fonte de ocorrência igual a u (u é a proporção de genes A1 que sofrem mutação para A2 entre uma geração e a seguinte). Se a frequência de variabilidade genética ao longo das cadeias evolutivas das espécies.
Do ponto de vista zootécnico, o efeito das mutações é praticamente desprezível no sentido de causar alterações nas características produtivas dos animais. Para características qualitativas, entretanto, algumas mutações podem ser importantes, como no caso do gene que condiciona ausência de chifres, na raça nelore (surgindo o Nelore variedade mocha).
Finalmente, as mutações podem ser dominantes ou recessivas de acordo com a ação do gene mutado.
fatores que alteram as taxas normais de mutação
a) Radiações ionizantes - muito bem conhecidas e estudadas nas quebras cromossômicas. Causam também grandes aumentos nas taxas de mutação (ex.: acidente nuclear de Chernobyl).
b) Agentes químicos - alguns como o gás mostarda ou o etilnitrosouréia são potentes agentes mutagênicos. Várias outras drogas têm sido estudadas, inclusive psicotrópicos.
c) Fatores ambientais - nutrição, temperatura, radiações naturais, agentes biológicos etc.
Ponto de equiíbrio
Suponha que o gene A1 sofra mutação para A2 com uma taxa de ocorrência igual a u (u é a proporção de genes A1 que sofrem mutação para A2 entre uma geração e a seguinte). Se a frequência de A1 em uma dada geração é p0 , a frequência de A2 surgidos por mutação, na geração seguinte, será up0 . Assim, a frequência de A1 na geração seguinte será p0
- up0 e a mudança na frequência gênica será - up0 . Considere-se, entretanto, a mutação ocorrendo em ambas as direções, supondo os alelos A1 e A2 com frequências p0 e q0 . A1 sofre mutação para A2 a uma taxa “u” por geração e A2 sofre mutação reversa a uma taxa “v”. Após uma geração haverá um ganho de A2 igual a up0 devido à mutação em uma direção e haverá
umaperda igual a vq0 devido à mutação reversa, ou seja,


conceituação
A seleção pode ser definida como o processo no qual alguns indivíduos são escolhidos entre os membros de uma população para produzirem a geração seguinte (lasley,1978). Pode ser de dois tipos: natural e artificial.
Deve-se considerar o fato de que os indivíduos de uma geração diferem em viabilidade e em fertilidade e que assim, contribuem com números desiguais de descendentes para a geração seguinte. O número de descendentes com que os indivíduos contribuem caracteriza o seu valor adaptativo(“Fitness”). Se as diferenças de “valor adaptativo”são, de alguma forma, associadas com a presença ou ausência de determinado gene, então a seleção opera naquele gene.
Quando o gene é sujeito à seleção, sua frequência nos descendentes é diferente da frequência nos pais, uma vez que pais de diferentes genótipos transmitem seus genes de forma diferenciada para a próxima geração. Dessa forma a seleção produz alteração na frequência gênica e, consequentemente, na frequência genotípica. decorrentes de seleção. Deve-se considerar o efeito da dominância com relação ao “valor adaptativo”(fig. 1).
Torna-se mais conveniente pensar na seleção agindo contra determinado gene, na forma de eliminação seletiva de um ou de outro genótipo do qual faz parte o gene em questão. Isto pode ocorrer sob a forma de redução da viabilidade ou da fertilidade, incluindo aí, a capacidade de se acasalar. No ciclo vital, a seleção age inicialmente sobre a viabilidade e depois sobre a fertilidade. Nas deduções da mudança na frequência gênica, considera-se o estádio de zigoto de uma geração até o mesmo estádio na geração descendente.
A intensidade da seleção é expressa como “coeficiente de seleção”(s) o qual representa a redução proporcional na contribuição do genótipo selecionado contra (1-s). Isto expressa o “valor adaptativo” de um genótipo em relação ao outro. Por exemplo, supondo-se que o “coeficiente de seleção seja s = 0,1 , então o “valor adaptativo (1-s) é igual a 0,9, significando que para cada 100 zigotos produzidos, 90 são produzidos pelo genótipo selecionado contra. O “valor adaptativo”sendo definido, como contribuição proporcional de descendentes, deveria ser chamado de “valor adaptativo relativo”, entretanto, é referido apenas como “valor adaptativo”.
O “valor adaptativo de um genótipo em relação a um determinado locus, não é
necessariamente o mesmo em todos os indivíduos. Depende de circunstâncias ambientais na qual os indivíduos vivem e também das interações gênicas (dominância e epistasia).
Quando se atribui um “valor adaptativo”a um genótipo, esse valor refere-se ao “valor adaptativo médio” na população como um todo.
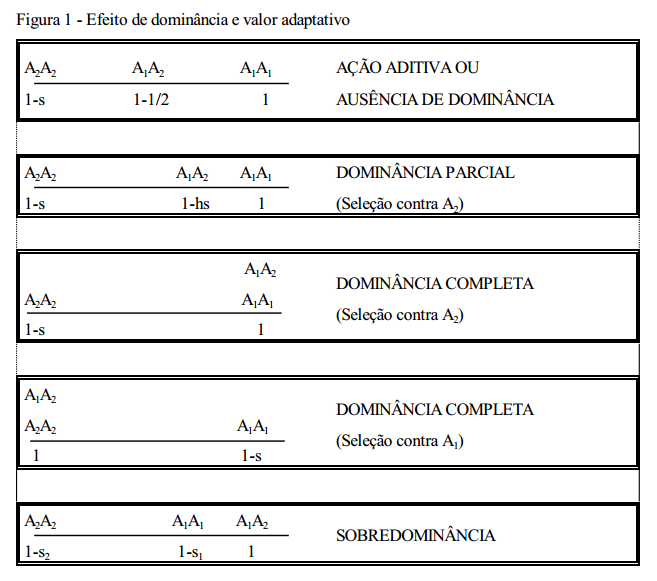
As deduções que se seguem, limitam-se a considerar os efeitos da seleção em um locus, muito embora as diferenças de “valor adaptativo” entre os indivíduos resultam da seleção atuando em muitos (quem sabe todos) os loci simultaneamente. Supõe-se, entretanto, que o “valor adaptativo médio” dos diferentes genótipos permaneçam constantes.
7 Fatores capazes de alterar as frequências gênicas II
Na dedução da equação de mudança da frequência gênica, as condições de dominância devem ser consideradas, mas o método é o mesmo em todas elas. Assim, pode-se deduzir uma fórmula básica para a variação da frequência gênica em uma geração de seleção.
Considere-se, como ilustração, o caso de dominância completa com a seleção agindo contra o homozigoto recessivo (Tabela abaixo):
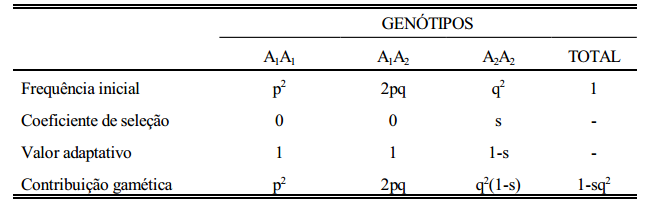
A primeira linha da tabela acima refere-se às frequências genotípicas antes da seleção, de acordo com o teorema de Hardy-Wenberg. Na segunda linha, s é o coeficiente de seleção contra o homozigoto recessivo (A2A2 ). Os valores adaptativos são: 1 (A1A1 ), 1 (A1A2 ) e 1-s (A2A2 ). Multiplicando-se a frequência inicial de cada genótipo pelo seu recessivo “valor adaptativo”, obtém-se a frequência do genótipo após a seleção (contribuição gamética). Após a seleção, a soma das frequências não é mais igual a 1, por causa da perda igual a sq 2 devida
à seleção contra o genótipo A2A2 . A frequência gênica após a seleção é calculada pelo método já visto no capítulo de cálculo de frequências gênicas.

O efeito da seleção na frequência gênica depende, portanto, da intensidade da seleção (s) e da frequência do gene na população. Para todas as condições de dominância, a dedução das equações segue o mesmo raciocínio. As equações são:


As conclusões acima aplicam-se tanto para a seleção natural quanto para a seleção artificial. A seleção artificial tem sido utilizada, de forma intuitiva, há milênios. Animais considerados superiores ou mais interessantes segundo algum critério, são intensamente utilizados na reprodução, alterando fortemente o equilíbrio das populações (equilíbrio de Hardy-Wenberg).
A seleção artificial será discutida mais detalhadamente no segundo volume deste material (Métodos Seleção)
oscilação genética
Também conhecida como “deriva genética”, é um processo dispersivo de alteração da frequência gênica. Ocorre em populações pequenas por “erro de amostragem”. Maiores detalhes poderão ser obtidos em strickberger, 1985 página 702-706.